Steinernema tami
Luc, Nguyen, Reid & Spiridonov, 2000
Description:
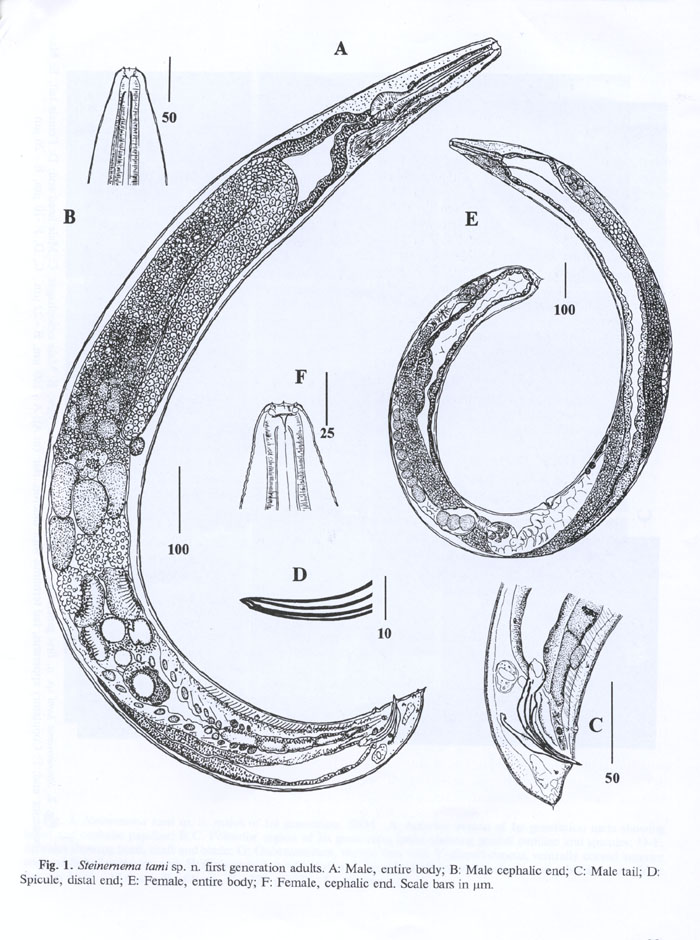
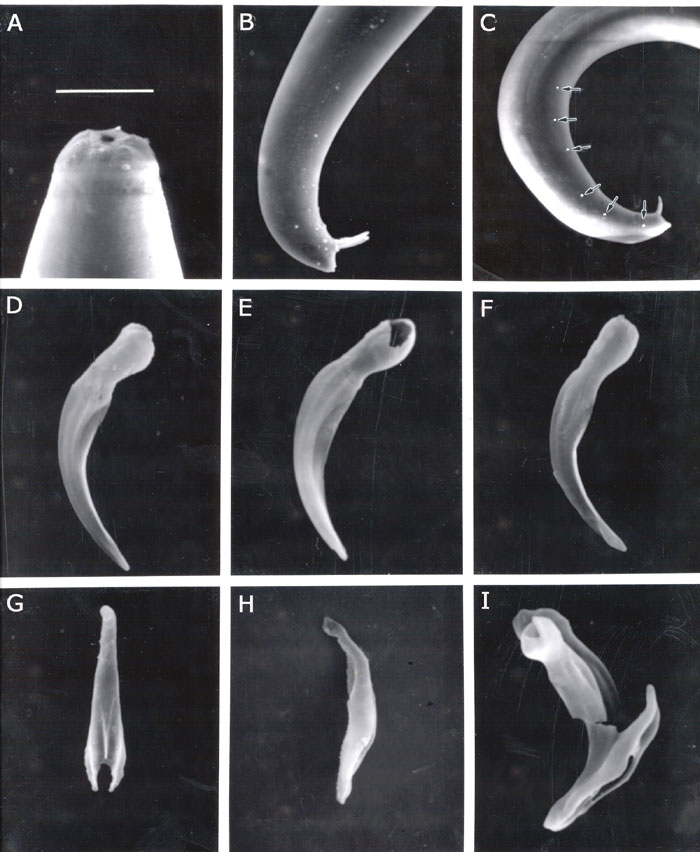
First generation males. (FIG. 1) Body slightly swollen in the middle. Heat-killed specimens C-shaped or J-shaped. Cuticle with faint transverse annulation poorly seen under light microscope. Head rounded with six low lips, 6 labial papillae (Fig. 1B) and four prominent cephalic papillae. Mouth opening more or less circular. Stoma about 5-6 mm deep with two sclerotized rings. Anterior ring lightly sclerotized, almost rounded, posterior ring more sclerotized, smaller, elliptical in shape and located on frontal border of oesophageal tissue. Oesophagus with procorpus, slightly swollen metacorpus, indistinct isthmus and pyriform basal bulb. Nerve ring surrounding isthmus. Cardia prominent. Excretory pore located slightly anterior to mid-esophagus, excretory duct about 20 mm long, cuticularized . Excretory glands swollen, 12-18 mm wide sometimes displacing basal bulb to dorsal side. Pseudocoelom in esophageal region filled with granular material. Distance from anterior end to testis flexure ranging from 280 to 350 mm. Different spermatogenesis stages visible in testis (Fig. 1A). In mid-testis region primarily small spermatocytes disappeared and replaced by about 30 (40-50 mm) large cells, variable in appearance, and often with wart-like structure on cell membrane . Closer to the vas deferens these structures appear nearly separated from the membrane, producing groups of refractive granules around large cells . Vas deferens is tightly filled with both large cells and granules. Spicules curved, yellow-brownish in color. Oblique muscles are well developed in precloacal zone (Fig. 1C). Spicule head (manubrium) length about 1- 1.5 times as long as wide, rarely twice (FIG. SEM1) . Spicule shaft (calomus) prominent. Blade (lamina) moderately curved bearing ribs reaching spicule tip. Velum thin. Spicula distal end bluntly pointed. Gubernaculum boat-shaped, proximal end slightly curved (Fig. 1C), in ventral view, corpus tapering gradually anteriorly, cuneus Y-shaped. Eleven pairs and a single genital papillae present, 6 pairs subventral, precloacal, a single larger precloacal midventral papilla, one pair sublateral precloacal, at level of mid lamina, one pair subventral adcloacal, two pairs subventral postcloacal, and one pair subdorsal postcloacal. Tail dorsally convex, conoid with a bluntly rounded terminus. Mucron 3-4 mm sometimes not observed. Phasmids inconspicuous.
Measurements. Holotype (Male, first generation): L = 1377 mm; body width =125 mm; distance from anterior end to excretory pore = 63 mm; total oesophagus = 153 mm; anal body width=38 mm; tail length =26 mm; spicule (chord) = 75 mm; spicule (arc) = 82 mm ; gubernaculum = 52 mm ; a = 11.0; b = 9.0; c =53.0; D= 0.4; E=2.4, SW = 1.97.
Paratype measurements: (Male, first generation, n=20). L= 1.600 (1.2-1.9) mm; W=129 (98-161) mm; EP=68 (43-92); ES=153 (137-166) mm; anal body width=37 (25-40) mm; tail = 23 (10-32) mm; spicule length along the chord = 72 (63-78) mm; spicule length along the arc = 77 (71-84) mm; D%= 44 (30-60); SW = 2.0 (1.4-3.0)
Second generation males. Heat killed specimens C-shaped. Anterior region similar to that of first generation males, but smaller, lips relatively more prominent. Excretory duct cuticularization, discernible only up to 3-4 mm from excretory pore. Testis reflexion usually behind mid body. Spermatogenesis is similar to that of first generation males, with up to 18 large cells with granules on the surface in vas deferens. Spicules and gubernaculum of the same color and shape as in first generation males. Distribution of genital papillae resembles that in first generation males, but very often postcloacal subventral papillae are displaced on the tail terminus with minute 2-3 mm long mucron protruding between papillae of posteriormost pair.
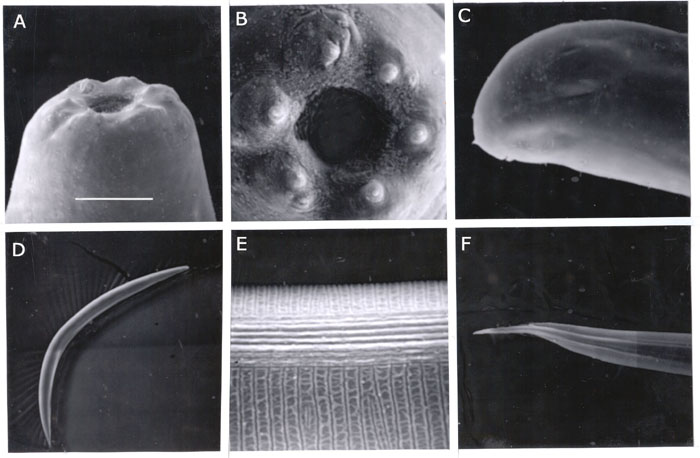
First generation females. Body robust, obese, usually C-shaped on heat relaxation. Anterior end truncate to rounded (FIG. SEM2) . Face view with rounded mouth, six labial papillae, and four cephalic papillae immediately posterior. Cervical part of body with coarse annulation. Lateral field inconspicuous. Stoma about 10-15 mm long and 8-10 mm wide, with two cuticular rings in the walls: anterior ring thick, and posterior one smaller, more refractive located on the esophageal tissue (Fig. 1F). Oesophagus more or less cylindrical with isthmus. Cardia prominent, well protruding into intestine lumen. Excretory pore anterior to mid esophagus. Excretory duct heavily cuticularized up to 30 mm from pore. Excretory gland large, elongated. Anterior ovary flexure situated about 100-200 mm posterior to basal bulb. About 10-20 amoeboid 20-25 x 30-45 mm spermatozoa in gonad lumen between ovary and uterus. Vagina short, sclerotized. Vulvar lips usually not protruding. Developing oocytes in multiple rows. Tail short, convex-conoid with a peg-like mucron.
Second generation females. General morphology of anterior end similar to that of first generation females. Excretory duct cuticularization visible up to 5-8 mm posterior to excretory pore. Cardia relatively more developed than in first generation females, especially when gonads less developed. Spermatozoa found in oviduct having the same morphology as in first generation females. Tail relatively longer than that in first generation females, conoid, terminus with a 2 mm long spike-like mucron. Posterior- anal swelling prominent.
Third stage infective juvenile (IJ3). Heat relaxed specimens ventrally arcuate . Body smoothly tapering towards both ends. Cephalic end rounded . Cuticle marked with prominent transverse annulation approximately one mm wide at midbody). Lateral fields at midbody with 8 ridges, 9 incisures) (FIG. SEM2) . At mid-body, central ridges are more elevated than marginal ones . On tail, most ridges disappeared gradually and the submarginal ones become prominent. Oesophagus narrow with a slender isthmus. Basal bulb subpyriform, usually displaced towards dorsal side by overdeveloped excretory gland. Hemizonid distinct, located in isthmus level. Excretory pore cuticularized, 2 mm in diameter in lateral view. Excretory duct less cuticularized than pore. Bacterial pouch rarely seen. Xenorhabdus cells, when found, with poorly visible walls, tightly compressed in bacterial chamber. Tail conoid, tapering to a finely pointed terminus. No refractive inclusion in the tip. Hyaline portion well pronounced, occupying about 39% of tail length. Phasmids distinct, located in anterior half of tail.
Measurements (n=20, all measurements are in micrometers): L = 530 (400-600) ; W = 23 (19-29); EP = 36 (34-41); ES = 117 (110-123); ABW = 12 (11-13); tail = 50 (42-57); a = 23 (19-28); b = 5 (3.7-5.1); c = 11 (9-12); hyaline part/tail = 0.4 (0.3-0.5); E% = 73 (67-86); D% = 31 (28-34).
Type host and locality. Natural hosts are unknown. The type locality is the primary forest in Cat Tien National Park along the Dong Nai river, Vietnam. The nematode was recovered by Galleria baiting using soil samples collected on 6-12 April 1997.
Type specimens. Holotype male (first generation) and numerous paratypes are deposited in the Collection of Invertebrates of the Institute of Parasitology, Russian Academy of Sciences, Moscow. Slides with one male and one female of first generation each are deposited in the Hanoi Institute of Ecology and Biological Resources, Vietnam; German Nematode Collection, Institute of Nematology & Vertebrate Studies, Munster, Germany; Collection of the International Institute of Parasitology (CABI Bioscience, Egham), England. Living cultures are maintained in Hanoi, Moscow and Egham.
Biology. Optimal temperatures for the infection of Galleria mellonella with infective juveniles of Steinernema tami sp. n. were between 22 and 27 o C. Between 22 and 24 o C the first generation males and females were observed inside Galleria cadavers on day 3 or 4 after inoculation . First generation adults disappeared on day 6 or 7, and numerous feeding juvenile stages of second generation were found in Galleria bodies. Early males and females of the 2nd generation were present in cadavers on day 8 or 9. Mass migration of the infective juveniles occured on day 13-16 depending on humidity. Prolonged storage at 3-5 o C increases juvenile mortality drastically.
Molecular diagnosis. The RFLP patterns yielded from each of six species (S. tami, S. carpocapsae, S. siamkayai, Malaysia , SSL1 and SSL 2) with seventeen different restriction enzymes showed that, S. tami is different from other species but molecularly closely related to them.
Differential diagnosis. Molecular
data indicates that Steinernema tami belongs to the group
of species related to Steinernema carpocapsae, namely the
"carpocapsae group" (body length <600 mm).
The new species can be distinguished from other species of the group by
the average body length of infective juveniles: S. tami, 530 (400-550)
mm,
S. kushidai (589 mm),
S. scapterisci
(572 mm), S. carpocapsae (558 mm),
S. abbasi (541 mm); but longer than that
of S. rarum, S. ritteri, and S. siamkayai (511, 510,
446 mm, respectively). Compared to
Steinernema
siamkayai:
S. siamkayai : tail=35.5 (31-41) mm
; EP = 57 (48-67) mm; E% = 96 (85-112)
S. tami : tail=50 (42-57)mm;
EP = 36 (34-41) mm; E% = 73 (67-86)
The D% distinguishes the S. tami (D% = 31(28-34) from
other steinernematids with short infective stage: S. abbasi, D%=53
(51-58), S. riobrave D% = 49 (45-55), and S. ritteri D% =
46 (44-50). Infective juveniles of S. tami is quite
different from that of S. kushidai but close to S. carpocapsae
and S. scapterisci. The new species can be separated from S.
carpocapsae and S. scapterisci by SEM of spicules and gubernacula
of these two nematodes (Hominick et al. 1997, Fig. 4) and those of S.
tami (in the reference at the end, Fig. 3).
Steinernema tami is also different from other
species in the group by male characteristics. Except for S. scapterisci,
spicule length (SL) = 83 mm, and S. siamkayai,
SL=75.5 mm, spicule length of S. tami, 72
mm
is longer than that of all other species in the "carpocapsae group": S.
ritteri, 69 mm;
S. carpocapsae, 66
mm; S. abbasi, 65 mm;
S kushidai, 63 mm;
S. rarum, 47
mm. The ratio SW also can be used to distinguish
S. tami from other species. SW of S. tami (SW=2.0), smaller
than that of S. scapterisci (SW=2.5) but larger than that of all
other species of the group: S. carpocapsae with SW=1.7; S. siamkayai,
SW=1.7; S. abbasi, SW=1.6; S. ritteri, SW=1.6; S. kushidai,
SW=1.5, and S. rarum, SW=0.94 (Nguyen & Smart, 1997).
The new species can be also separated from closely related species
by rDNA-RFLP analysis. RFLP profiles show that Steinernema tami differs
from other species of this group in the profiles of at least 3-4 enzymes.
This document was constructed and is maintained by KHUONG
B. NGUYEN
Entomology & Nematology Department
University of Florida
{kind=link}
{kind=link}
{kind=link}