Steinernema
beddingi Qiu, Hu, Zhou, Pang & Nguyen, 2005
Description
Male, first generation:
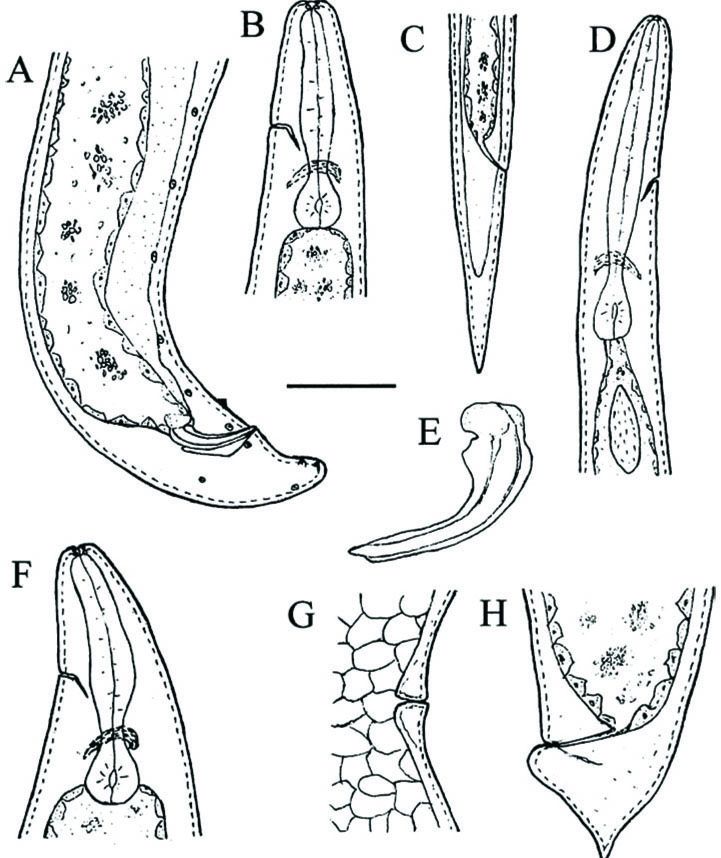
(Fig. 1) Body C-shaped and curved
posteriorly when killed by gentle heat. Cuticle smooth. Stoma shallow and wide.
Pharynx extending nearly to mouth opening. Cheilorhabdions and prorhabdions
sclerotised, small but distinct. Pharynx muscular with a cylindrical procorpus,
metacorpus swollen, isthmus well defined, basal bulb enlarged. Nerve ring often
surrounding anterior portion of basal bulb. Excretory pore opening located near
posterior part of metacorpus. Lateral fields and phasmids not observed. Gonad
single and reflexed. Spicules paired, colorless and moderately curved. Spicule
length/width about 4.6. Spicule head shorter than wide with a length/width
about 1:2 (Fig. 3). Lateral lobe prominent,
beginning anteriorly at shaft and extending posteriorly to spicule tip. Ventral
lobe enlarged anteriorly and ventrally to form a prominent rostrum and ending
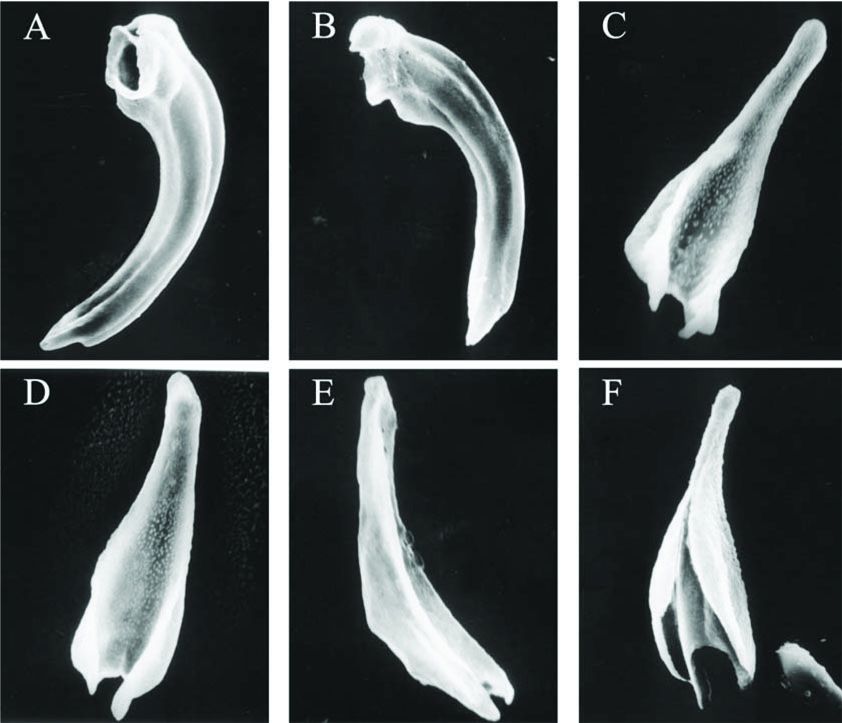
before, but close to, spicule tip (Fig. 1E; Fig. 4A and B). Dorsal lob starting
from shaft and terminating at a level similar to ventral lob. Spicule terminus
blunt. Velum not observed. Gubernaculum boat-shaped in lateral view. In ventral
view, gubernaculum tapering anteriorly and evenly. Corpus fused and cuneus not
seen (Fig. 4C to F). Tail long and concave ventrally. No mucron-like projection
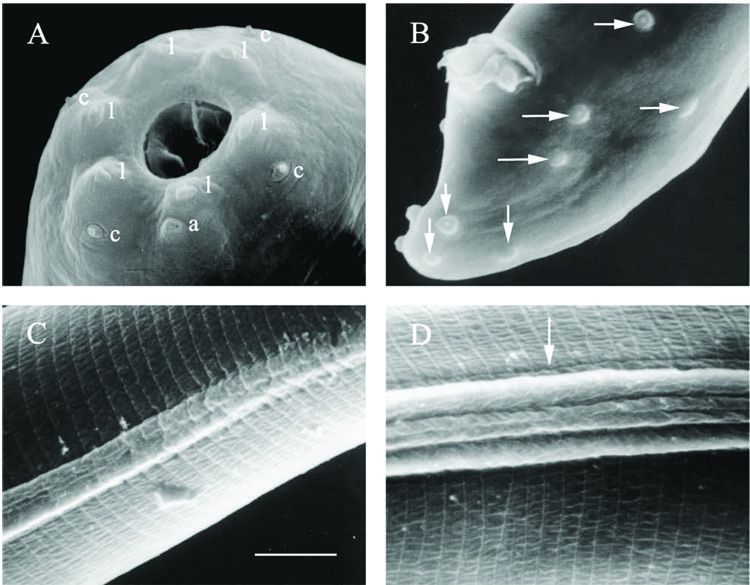
was observed at tip of tail. Twenty-three genital papillae were seen by SEM,
including five pairs and one single preanal papillae, two pairs adanal, one
pair lateral and three pair postanal (two subterminal and one subdorsal)
papillae (Fig. 1B; Fig 3F).
Male,
second generation: Morphologically similar to
the first generation male, such as tail concaves ventrally and without mucron,
but most of measurements are smaller and excretory pore is located more
anterior than that in the first generation.
Female, first
generation: (Fig. 2) Body C-shaped when
killed by gentle heat. Cuticle smooth, head rounded and slightly truncated
anteriorly. Six lips fused, each with a papilla. Four cephalic papillae occur
further back on head. Amphidial aperture distinct and located behind lateral
labial papillae. Morphology of anterior region of female similar to male.
Lateral fields and phasmids not observed. Gonads didelphic and reflexed. Vulva
a transverse slit, usually slightly protruding and symmetrical. Vagina short
and muscular. Epiptygma not observed. Tail wide and with a wedge-like
projection on tip. Postanal swelling distinct.
Female, second
generation: Similar to the first generation females but much smaller. Vulva
is slightly more protruding than the first generation. Postanal swelling and
tail projection as prominent as those of the first generation.
Infective juveniles: Body
slender, enclosed in a sheath. Anterior end rounded, labial region smooth,
papillae not seen. Amphidian apertures slit�Cshaped. Four prominent cephalic
papillae. Mouth and anus closed. Pharynx long and narrow, isthmus distinct and
surrounded by nerve ring. Excretory pore located anterior to nerve ring, near
middle of pharynx. Cuticle with prominent transverse striations. Lateral field
begining anteriorly with two slightly raised ridges at a position slightly
anterior to excretory pore. Another two ridges appear between previous two at a
position close to base of pharynx and become gradually more prominent at middle
portion of body and then fade gradually after level of anus (Fig. 3D and E).
Middle two ridges slightly smaller than other two and whole lateral field
distinctly protrudes from body surface. Tail attenuate and tapering gradually
with a hyaline portion occupying about 45% of tail length. No spine like
structure was seen on tail tip. Phasmid inconspicuous. There was no substantial
difference between IJ produced from the artificial medium and from Galleria
larvae except that the former was slightly short than the later (Table 1).
S.
beddingi is
characterised morphologically by the combination of the features of various
developmental stages of the nematode. For IJ, body length averaging 743 (700-790) ��m; distance from anterior end to excretory pore 70 (64-75) ��m, to the base of pharynx 125
(113-130) ��m; tail length 77
(72-83) ��m; D and E value 57 (52-64)
and 92 (84-103), respectively; lateral field at the mid-body position
has only four ridges, which raise distinctly from body surface; tail attenuates
and tapering gradually without a dorsal constrict or a spine on the tail tip.
For males, tail has a concave in the ventral part and without mucron on tip;
spicules and gubernacula are colorless; spicule head very short, dorsal and
ventral lobs ending before the spicule tip while distal end of lateral lob
suddenly narrowed both dorsally and ventrally to form a blunt tip; gubernaculum
without cuneus. For females, vulva is a symmetric, transverse-slit without
epiptygma, tail with a wedge-like structure on the tip and a conspicuous
postanal swelling.
The sequence length and composition of ITS
regions (AY603397) of S. beddingi are 1069 bp
and 280A, 234G, 373T, 182C, GC% = 38.9, and partial 28S rDNA (AY603396)
are 465 bp and 123A, 138G, 127T,
77C, GC% = 46.2.
Both
morphometrical (Tables 1) and molecular (Fig. 5-8) data show congruently that S.
beddingi n. sp. is close to S. affine and S. intermedium.
However, the new species can be separated from these two described species by
the morphological characters of various developmental stages of the nematodes
(Table 4). The morphometrics of S. beddingi IJ are very close to those
of S. affine and S. intermedium except that the tail is longer,
77 (72-83) ��m compared to 64 (53-72) ��m for S. intermedium
and 66 (64-74) ��m for S. affine. IJ of the new species can also
be distinguished from the two described species mentioned above by morphology
of tail and lateral field: S.
beddingi IJ tail attenuates
and tapering gradually with no spine on tail tip, while S. affine IJ
usually has a spine like structure on tail tip, and tail of S. intermedium
IJ usually has a constriction on the dorsal side. The middle portion of lateral
field of the new species has four raised ridges while that of the other two
species has 6 ridges. For the first generation males, the spicule and
gubernaculum length of S. beddingi, 71 (63-78) ��m and 43 (38-48)
��m, respectively, is shorter than those of S. intermedium 91
(84-100) ��m and 64 (56-75) ��m, respectively. The morphology of
spicule and gubernaculum also differed substantially from the two described
species (Adams & Nguyen 2002; Fig. 4), for example, no velum was observed
on spicules of S. beddingi n. sp while it was observed clearly on
spicules of both S. intermedium and S. affine; no cuneus was
observed on S. beddingi gubernacula while both S. affine and
S. intermedium has a needle type cuneus. For the first generation females,
the new species has a distinct postanal swelling but the other two species have
not. S. beddingi can also
be distinguished from S. affine and S. intermedium genetically by
either the ITS or 28S D3 domain rDNA sequence, and biologically by cross tests.
S. beddingi can be distinguished from
other described Steinernema species that have a similar mean IJ body
length (ranged from 650 ��m to
850 ��m), including S.
pakistanense Shahina, Anis, Reid, Rowe and Maqbool 2001, S. ceratophorum
Jian, Reid and Hunt 1997, S. bicornutum Tallosi, Peters, Ehlers
1995, S. feltiae (Filipjev, 1934) Wouts, Mracek, Gerdin and Bedding
1982, S. thanhi Phan, Nguyen and Moens 2001, S. sangi Phan,
Nguyen and Moens 2001, S. weiseri Mracek, Sturhan and Reid 2003 and S.
monticolum Stock, Choo and Kaya 1997 by morphological characters of
various stages. For example, it can be distinguished from all nematodes
mentioned above by the spicule and gubernaculum morphology of the first
generation males (Fig. 4; Adams and Nguyen, 2002) as well as lateral field
morphology and ridge number of IJ (the number of ridges at the middle portion
of the above mentioned nematodes is six to eight while that of the new species
is four and the whole lateral filed raised distinctly from body surface). The
new species can also be separated from S. pakistanense, S. ceratophorum and
S. bicornutum by IJ head structure (IJ of these three species possess a
horn like structure on their head).
Type host and locality
The type host of this
nematode in nature is unknown as it was recovered from soil using Galleria
larvae as bait. The soil sample was collected from a cabbage field in
An-fen-ying town (latitude N24.58,
longitude E102.28, altitude 1850m, mean annual temperature 16oC,
precipitation 1000 mm/year), about 60 km from Kunming, Yunnan province, People��s Republic of
China.
Type materials
Holotype male, allotype
female, five paratype first generation males, five paratype females, fifteen
paratype infective juveniles and other population slides deposited in the State
Key Lab for Biocontrol, School of Life Sciences, Zhongshan University,
Guangzhou 510275, China. One slide that contains about ten paratype IJ and two
slides, each contains one paratype male and one female will be deposited in
USDA Nematode Collection, Beltsville, Maryland, USA. Living infective juveniles
is also preserved in liquid nitrogen in the nematode collection of SKLB,
Zhongshan University.
S. beddingi reproduced
poorly in Galleria larvae, usually
without second generation adults in the infected Galleria
larval cadaver and only a few thousands IJ were obtained from a larvae
averagely. However, it proliferated well in the artificial medium using the
method described. IJ produced either from Galleria
larvae or the artificial medium were highly pathogenic to Galleria larvae, with most of larvae dying within 24
hrs when 15 larvae were exposed to about 2000 IJ in a Petri dish lined with two
layer of moist filter paper at 23 oC. It is unknown why S. beddingi n. sp. reproduced poorly in Galleria larvae. The symbiotic bacteria of S. beddingi n. sp. have been isolated and preserved in
liquid nitrogen. It produced a yellow pigment on the artificial medium
mentioned and possessed
many characters of Xenorhabdus species, such as phase variation,
negative to peroxide, etc. A phylogenetic tree based on the 16S rDNA sequence
showed the symbiotic bacteria of S. beddingi n. sp. claded with but can
be clearly distinguished from X. bovienii (Qiu et al.,
unpublished data).
Etymology
The
species was named after R. A Bedding from the Australian Center for International Agricultural Research (ACIAR).
Qiu, L, Hu, X.., Zhou, Y., Pang, Y.
and Nguyen K. B. 2005.
Steinernema beddingi n. sp. (Nematoda:Steinernematidae), a new
entomopathogenic nematodes fron Yunan, China. Nematology 7, 737-749.
{kind=link}
{kind=link}
{kind=link}