common name: wedge-shaped beetles (suggested common name)
scientific name: Ripiphorus spp. (Insecta: Coleoptera: Ripiphoridae)
Introduction - Distribution - Description - Life Cycle - Hosts - Economic Impact - Selected References
Introduction (Back to Top)
Ripiphoridae are a family of unusual parasitic beetles that are thought to be related to tumbling flower beetles (Coleoptera: Mordellidae) and blister beetles (Coleoptera: Meloidae). There is disagreement over the spelling of the family (Ripiphoridae) and genus (Ripiphorus) names. Here we use the original spelling that starts with only the letter “R”; however, an initial “Rh” has also been used in the scientific community (Rhipiphoridae and Rhipiphorus).
Generally, the biology of the family Ripiphoridae is poorly known. Ripiphorids parasitize bees and wasps (Hymenoptera), roaches (Blattodea), and wood-boring beetles (Coleoptera). However, the specific hosts for many ripiphorid species are unknown. Furthermore, only one sex (either male or female) has been described for several species, and the males and females of some species look different.
Two genera of Ripiphoridae infest hymenopteran (bee and wasp) nests: Macrosiagon Hentz (Figure 1A) and Ripiphorus Bosc(formerly Myodites Latreille) (Figure 1B). Species of Macrosiagon are parasites of a variety of Hymenopteran families including: Halictidae, Vespidae, Tiphiidae, Apidae, Pompilidae, Crabronidae, and Sphecidae. Ripiphorus species parasitize only bees in the families Halictidae and Apidae.
Figure 1. Adult specimens of the two genera of Ripiphoridae. A) Macrosiagon Hentz, and B) Ripiphorus Bosc. Photographs by Allen M. Boatman.

There are an estimated 35 nearctic species of Ripiphorus, two of which have been collected in Florida: Ripiphorus schwarzi LeConte (Figure 2A) and Ripiphorus fasciatus Say (Figure 2B). Due to limited information for both of these species, the information presented below is characteristic of the genus Ripiphorus. Information specific to Ripiphorus fasciatus and Ripiphorus schwarzi is presented where detailed information is available.
Figure 2. Adult specimens of the two species of Ripiphorus present in Florida, A) Ripiphorus schwarzi LeConte and B) Ripiphorus fasciatus Say. Photographs by Tom Murray. Licensed under Creative Commons Attribution 1.0 via bugguide.net.

Distribution (Back to Top)
Ripiphorus species occur worldwide, except in Australia. They are distributed widely across North America, but they are most diverse in the southwestern United States deserts. Ripiphorus fasciatus has been collected from Atlantic Canada to the north, Kansas to the west, and Florida to the south. Ripiphorus schwarzi has been observed in Massachusetts, Virginia, and Florida.
Description (Back to Top)
Ripiphorus species are holometabolous (undergo complete metamorphosis). Descriptions of the egg, larval, and pupal stages are available only for one species, Ripiphorus smithi Linsley & MacSwain. Therefore, the information provided in the sections below is a generalization based on what is known about that species. A description of adult Ripiphorus fasciatus is available and is described in the “Adults” section below.
Eggs: Eggs are typically 0.5 mm long, 0.2 mm wide, and tapered.
Larvae: First instar larvae are heavily sclerotized, highly mobile, and are referred to as triungulins (Figure 3A). They are equipped with an anal sucker and have a large sucker type pad on the ends of the tarsi. The second through sixth larval instars are morphologically very different from the first larval instar. The later instars are much less mobile, unsclerotized (soft-bodied), and have large tubercles (round projections) present on each segment (Figure 3B).
Figure 3. A) First and B) sixth (final) Ripiphorus larval instars. Note that the sixth larval instar image is not to scale. Figure adapted, with editorial permission from Tomlin and Miller 1989 (A) and Linsley et al. 1952 (B).
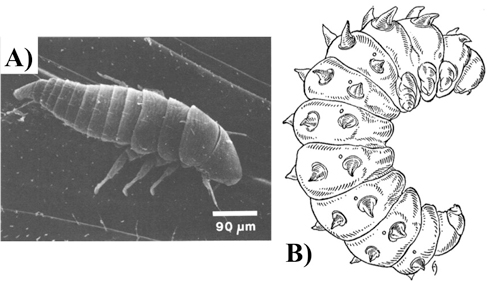
Pupae: The structural form of the pupae is generally quite similar to the adult form of the same sex. The primary exceptions to this are that the hind wings are not unfolded and the abdomen is almost completely free of hair in pupae.
Figure 4. Ventral view of a Ripiphorus pupa. Figure adapted, with editorial permission, from Linsley et al. 1952.
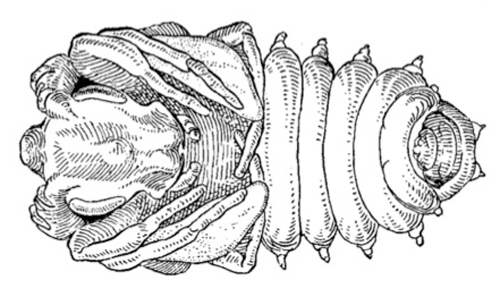
Adults: Ripiphorus fasciatus adults are 6-8 mm long and have a fly-like appearance because the elytra (1st pair of wings) are short, leaving the large, unfolded hind wings exposed. The hind wings are clear and the elytra are bicolored; the base of the hind wing is brown while the rest is yellow. The body is thick and appears humpbacked because the abdomen curls under the body. Their large eyes protrude from the head and the antennae consists of 10-11 segments. Males have bi-flabellate antennae (Fig. 5A), and females have pectinate antennae (Fig. 5B).
Figure 5. Male and female antennal types. A) Male, bi-flabellate antennae: note the protruding filaments from both sides of the central antennal stalk. B) Female, pectinate antennae: note that the filaments protrude only from 1 side of the central antennal stalk. Photographs by A) USGS Bee Inventory and Monitoring Lab. Licensed under Creative Commons Attribution 2.0 via flickr and B) Tom Murray. Licensed under Creative Commons Attribution 1.0 via bugguide.net.

Life Cycle (Back to Top)
Ripiphorus smithi is the only species of Ripiphorus for which a detailed life history is known. The life histories of all Ripiphorus species are presumed to be similar to that of Ripiphorus smithi. Therefore, the information provided in this section a generalization based on what is known about Ripiphorus smithi.
Adult Ripiphorus emerge during the summer. After mating, the females deposit 6 to 15 eggs into the buds of unopened flowers. The total number of eggs that each female lays is unknown, but dissected females have contained as many as 850+ eggs. When the flower opens, the eggs hatch and the first instar Ripiphorus begin to seek a host (Figure 6A). The larvae will “stand” on the tip of their abdomens until an adult bee of their host species lands on the flower. The larvae then attach themselves to the adult bee that carries them back to their nest.
The Ripiphorus larva enters the host bee’s nest and waits on the bee’s pollen mass until an egg is laid and hatches. The Ripiphorus larva penetrates the bee larva and overwinters in the diapausing bee larva (Figure 6B). When the bee reaches the pre-pupal stage, in the late spring/early summer, the Ripiphorus larva grows quickly and emerges from the thorax of the bee. As it emerges, the Ripiphorus larva molts and the molted exoskeleton plugs the wound on the honey bee pre-pupa (Figure 6C). The Ripiphorus larva continues to feed on the same bee through four more molts until the bee is consumed, at which point the Ripiphorus pupates for 11-14 days. Superparasitism of a single bee larva by multiple Ripiphorus is rare, and when it does occur, neither of the Ripiphorus larvae completes development.
Figure 6. First larval instar phases: A) dorsal and lateral view of the host-seeking phase, B) lateral view of the overwintering endoparasitic phase, C) lateral view of the final endoparasitic phase- immediately prior to emerging from the bee pre-pupa and molting to the second instar. Figure adapted, with editorial permission, from Linsley et al. 1952.
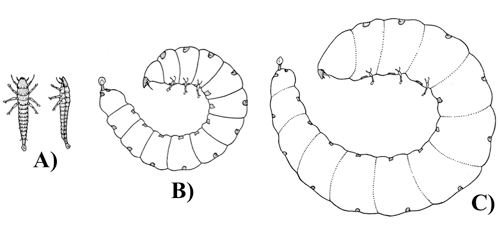
The adult life stage is very short (as few as 1-2 days; Table 1). After emergence, male Ripiphorus fly over the bee’s nesting areas to locate females for mating. It is suspected that the females release a strong sex pheromone that aids in the attraction of mates. Males may mate with two or more females, but females only mate once. Multiple males typically swarm to a single female. After the female has successfully copulated, the males disperse to find unmated females.
Table 1. Seasonality of Ripiphorus life stages. Table adapted from Linsley et al. 1952.
Hosts (Back to Top)
Ripiphorus species parasitize two families of bees, Apidae and Halictidae. Within Apidae, one genus, Diadasia Patton, is parasitized. Several genera within Halictidae (Dieunomia Cockerell, Halictus Latreille, Lasioglossum Curtis, Augochlora Smith, and Augochlorella Sandhouse) are parasitized.
Ripiphorus fasciatus parasitizes Lasioglossum Curtis and, possibly, Halictus Latreille species (Figure 7A & B). Ripiphorus schwarzi also parasitizes Lasioglossum bees and has been reported to parasitize a species of Augochlora (Figure 7B & C). However, there is some debate that the record on Augochlora pura Say may have been a misidentification of another species of Ripiphorus.
Figure 7. Examples of Ripiphorus fasciatus and Ripiphorus schwarzi host species. A) Lasioglossum spp. is parasitized by Ripiphorus fasciatus and Ripiphorus schwarzi. B) Halictus spp. maybe parasitized by Ripiphorus fasciatus. C) Augochlora pura maybe parasitized by Ripiphorus schwarzi. Photographs by A) David Cappaert, Michigan State University, Bugwood.org; B) Joseph Berger, Bugwood.org; C) The Packer Lab - Bee Tribes of the World, via Wikimedia Commons.

Economic Importance (Back to Top)
Ripiphorus is distributed widely and can be locally abundant. However, their densities vary regionally and the reported rates of Ripiphorus infestation range from <1% to 28%. Ripiphorus are seldom encountered in nature because their immature stages occur primarily within the host’s nest and their free-living adult stage is very short (1-2 days). Their secretive life cycle makes an assessment of their economic and ecological impact very difficult. Additional research is necessary to determine the abundance and impact of Ripiphorus species.
Selected References (Back to Top)
- Batelka J, Hoehn P. 2007. Report on the host associations of the genus Macrosiagon (Coleoptera: Ripiphoridae) in Sulawesi (Indonesia). Acta Entomologica Musei Nationalis Pragae 47: 143-152.
- Batra SWT. 1965. Organisms associated with Lasioglossum zephyrum (Hymenoptera: Halictidae). Journal of the Kansas Entomological Society 38: 367-389.
- Buckley K, Nalen CZ, Ellis J. Halictidae (Insecta: Hymenoptera: Halictidae). University of Florida, IFAS, Entomology and Nematology Department, Featured Creatures, EENY-499. (2011). (28 October 2014).
- Eggleton P, Belshaw R. 1992. Insect parasitoids: An evolutionary overview. Philosophical Transactions of the Royal Society of London: Biological Sciences 337: 1-20. doi: http://dx.doi.org/10.1098/rstb.1992.0079.
- Falin ZH. 2002. Ripiphoridae Gemminger and Harold 1870 (1853) pp. 431-444 in Arnett RH, Thomas MC, Skelley PE, Frank JH. American Beetles. Volume 2, Polyphaga: Scarabaeoidea through Curculionoidea. CRC Press, Boca Raton.
- Falin ZH, Areneson LC, Wcislo WT. 2000. Night-flying sweat bees Megalopta genalis and M. ecuadoria (Hymenoptera: Halictidae) as hosts of the parasitoid beetle Macrosiagon gracilis (Coleoptera: Rhipiphoridae). Journal of the Kansas Entomological Society 73: 183-185.
- Linsley EG, MacSwain JW, Smith RF. 1952. The life history and development of Rhipiphorus smithi with notes on their phylogenetic significance (Coleoptera: Rhipiphoridae). University of California Publications in Entomology 9: 291-314.
- Majka CG, Chandler DS, Sheffield CS, Webster RP. 2006. New records of Ripiphoridae (Coleoptera) from the Maritime Provinces of Canada. The Coleopterists Bulletin 60: 299-303. doi:
- Peck SB, Thomas MC. 1998. A distributional checklist of the beetles (Coleoptera) of Florida. Florida State Collection of Arthropods, Arthropods of Florida and neighboring land areas 16: 103. (1998). (28 October 2014).
- Rivnay E. 1929. Revision of the Rhipiphoridae of North and Central America (Coleoptera). Memoirs of the American Entomological Society 6: 68.
- Sikes DS. 2003. The beetle fauna of the state of Rhode Island, USA (Coleoptera): 656 new state records. Zootaxa 340: 1-38.
- Tomlin AD, Miller JJ. 1989. Physical and behavioral factors governing the pattern and distribution of Rhipiphoridae (Coleoptera) attached to wings of Halictidae (Hymenoptera). Annals of the Entomological Society of America 82: 785-791.
- Vaurie P. 1955. A review of the genus Macrosiagon in Mexico, with notes on Rhipiphorus (Coleoptera, Rhipiphoridae). American Museum of Natural History, American Museum Novitates 1717. (28 October 2014).