common name: Polydamas swallowtail, gold rim swallowtail, tailless swallowtail
scientific name: Battus polydamas lucayus (Rothschild & Jordan) (Insecta: Lepidoptera: Papilionidae: Papilioninae: Troidini)
Introduction - Nomenclature - Distribution - Description - Life Cycle - Host Plants - Medicinal Uses - Defenses - Acknowledgements - Selected References
Introduction (Back to Top)
The Polydamas swallowtail is one of only two United States swallowtails of the genus Battus. It is our only United States swallowtail without tails (Figure 1).
Figure 1. Polydamas swallowtail, (Battus polydamas lucayus [Rothschild & Jordan]). Photograph by Donald W. Hall, University of Florida.
![Polydamas swallowtail, (Battus polydamas lucayus [Rothschild & Jordan])](Battus_polydamas01.jpg)
Nomenclature (Back to Top)
The Polydamas swallowtail was originally described and named Papilio polydamas by Linnaeus but was later transferred to the genus Battus (Scopoli 1777) and selected as type species of the genus by Lindsey (1925).
The name Battus is from Battus I, founder of the ancient Greek colony Cyrenaica and its capital, Cyrene, in Africa. The specific epithet polydamas is from Polydamas, a character from Greek mythology (Opler & Krizek 1984).
Larvae of the Polydamas swallowtail and those of the other swallowtails belonging to the tribe Troidini feed exclusively on plants in the genus Aristolochia and are commonly referred to as the Aristolochia swallowtails.
Distribution (Back to Top)
In the United States, Battus polydamas lucayus occurs as a regular resident in peninsular Florida and the Florida Keys, and occasional strays wander to other Gulf states and as far north as Missouri and Kentucky (Scott 1986). Subspecies polydamas occurs in southern Texas (Figure 2) (dos Passos 1940).
Figure 2. Polydamas swallowtail, (Battus polydamas lucayus [Rothschild & Jordan]), United States distribution map. Map by Donald W. Hall, University of Florida. (Based on dos Passos 1940, Minno et al. 2005, Opler & Krizek 1984, Scott 1986).
![Polydamas swallowtail, (Battus polydamas lucayus [Rothschild & Jordan]), United States distribution map](Battus_polydamas02.jpg)
Description (Back to Top)
Adults: The wingspread range is 9.2-11.6 cm. (approx. 3.6-4.6 in.) (Opler & Malikul 1998). Females are larger than males. The upper surface of the wings is black with submarginal yellow bands and marginal yellow spots. The underside of the wings is black with a submarginal row of yellow spots on the front wing and a sub-marginal row of wavy red lines and a marginal row of yellow spots on the hind wing. The body of the adult is black with red dots dorsally on the anterior part of the thorax and red-orange dots laterally on the thorax and coxae. A reddish-orange lateral line runs the length of the abdomen (Figure 3).
The yellow bands of lucayus are wider than those of subspecies polydamas, and the underside of the hindwing of subspecies lucayus is paler than that of subspecies polydamas (Rothschild & Jordan 1906).
Figure 3. Adult Polydamas swallowtail, (Battus polydamas lucayus [Rothschild & Jordan]), dorsal (top) and ventral (bottom) views. Photographs by Donald W. Hall, University of Florida.
![Adult Polydamas swallowtail, (Battus polydamas lucayus [Rothschild & Jordan]), dorsal (top) and ventral (bottom) views](Battus_polydamas03.jpg)
Battus males have androconia (scent scales) hidden in a fold of the inner margin on the upper surface of the hind wing (Field 1938) (Figure 4). The scales are fluted - possibly to increase the surface area for release of pheromones (Miller 1987, Racheli & Oliverio 1993). When courting, males “helicopter” around the females while fanning the androconial chemicals over them (Tyler et al. 1994, p. 53).
Figure 4. Male Polydamas swallowtail, (Battus polydamas lucayus [Rothschild & Jordan]), showing location of androconia (scent scales) along vein on the inner margin of the hind wing. Insets: magnified area of vein (lower left) and androconia (upper left). Photographs by Donald W. Hall, University of Florida.
![Male Polydamas swallowtail, (Battus polydamas lucayus [Rothschild & Jordan]), showing location of androconia (scent scales) along vein on the inner margin of the hind wing. Insets: magnified area of vein (lower left) and androconia (upper left)](Battus_polydamas04.jpg)
Eggs: Eggs are approximately 1 mm in diameter and vary from yellow to orange – appearing slightly greenish as they mature - due to the developing larvae that show through the egg shell (Figure 5). Eggs of our other U.S. Battus species (Battus philenor [L.]) are reddish-orange.
The eggs of all Aristolochia (troidine) swallowtails are partially covered by a hard, nutritious secretion that is usually laid down in vertical bands (Comstock & Grimshawe 1935, Tyler et al. 1994). There are numerous relatively large droplets on the bands. The secretion is produced by a large gland that lies above the female’s ovipository duct (Tyler et al. 1994).
Figure 5. Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]), eggs on leaf (left) and stem (right). Photographs by Donald W. Hall, University of Florida.
![Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]), eggs on leaf (left) and stem (right)](Battus_polydamas05.jpg)
Larvae: Full-grown larvae are approximately 5.3 cm (approximately 2.1 in) in length (Minno et al. 2005). Detailed descriptions of the larval instars were given by Brown et al. (1980[81]) and Comstock and Grimshawe (1935).
First instar larvae are tan and have numerous small tubercles – each bearing a single hair (Figure 6). After molting to the second instar, larvae are dark brown and have larger tubercles – each bearing numerous hairs (Figure 6).
Figure 6. Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]), first (bottom) and second (top) instar larvae. Photograph by Donald W. Hall, University of Florida.
![Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]), first (bottom) and second (top) instar larvae](Battus_polydamas06.jpg)
The cuticle of second instar larvae has a rough-textured appearance (Figure 6) whereas the cuticle of third (and subsequent) instars appears smoother and somewhat glossy (Figure 7).
Figure 7. Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]), third instar larvae. Note smooth, glossy cuticle. Photograph by Donald W. Hall, University of Florida.
![Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]), third instar larvae. Note smooth, glossy cuticle](Battus_polydamas07.jpg)
Full-grown larvae are variable in color - ranging from dark brown with black-tipped, orange tubercles to light tan with pale tubercles (Figure 8). The cuticle also has a pattern of darker lines that may be difficult to see in the darkest specimens (Figure 8, bottom image). Lighter colored specimens are less common.
Figure 8. Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]), full-grown larvae (fifth instars) showing some of the color variation. Photographs by Donald W. Hall, University of Florida.
![Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]), full-grown larvae (fifth instars) showing some of the color variation](Battus_polydamas08.jpg)
Pupae: Pupae may be either brown or green (Figure 9).
Figure 9. Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]), green and brown pupae. Photographs by Donald W. Hall, University of Florida.
![Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]), green and brown pupae](Battus_polydamas09.jpg)
Life Cycle (Back to Top)
Polydamas swallowtails have many flights all year in southern Florida and southern Texas, but they are rare during the winter northward. They favor open woods, gardens, and disturbed areas (Brown et al. 1980[81], Spade et al. 1988). Volatiles from the pipevine host plants may aid females in their search for oviposition sites (Pinto et al. 2009b). Eggs are laid preferentially on plants in sunny areas even though survival of larvae is greater in shady areas (Rausher 1979).
Eggs are laid in small groups on stems or less frequently on new leaves (Figure 5) of pipevines (Aristolochia: Aristolochiaceae). In Lepidoptera eggs, a small quantity of yolk remains trapped between two of the embryonic membranes (amniotic and serosa) that remain inside the egg shells after hatching (Barbier & Chauvin 1976, Richards & Davies 1977 [p.337]). Soon after hatching, larvae eat the egg shells (Figure 10), and the residual embryonic yolk and the secretion on the outside of the eggs (Moss 1919, Tyler et al. 1994, p. 113) serve as their first meal. Larvae also eat their exuviae (old exoskeleton) after each molt to conserve nutrients (Figure 11).
Figure 10. Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]), newly emerged larvae eating egg shells (chorions). Photograph by Donald W. Hall, University of Florida.
Figure 11. Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]). Fifth (last) instar larva immediately after molting (top) and eating exuviae (bottom). Photographs by Donald W. Hall, University of Florida.
![Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]), newly emerged larvae eating egg shells (chorions)](Battus_polydamas10.jpg)
![Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]). Fifth (last) instar larva immediately after molting (top) and eating exuviae (bottom)](Battus_polydamas11.jpg)
Young larvae feed on the tender new leaves and young stems. Older larvae will also eat the older leaves, stems, and flowers of the host plants. The aristolochic acids of the host plants may serve as feeding stimulants (Pinto et al. 2009a). Young larvae are gregarious (Figure 12), but become solitary as they mature. Sometimes hosts are completely defoliated, and larvae may find new hosts by detecting volatiles from nearby plants (Pinto et al. 2009b).
Figure 12. Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]). Gregarious third instar larvae. Photograph by Donald W. Hall, University of Florida.
![Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]). Gregarious third instar larvae](Battus_polydamas12.jpg)
Full-grown larvae typically migrate away from the host plant for pupation. In preparation for pupation, they attach by their terminal set of prolegs to silk pads spun on the new substrate and hang by silk girdles also attached to the substrate (Figure 13).
Figure 13. Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]). Prepupae clinging to silk pads with their terminal prolegs and hanging by silk girdles. Photograph by Donald W. Hall, University of Florida.
![Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]). Prepupae clinging to silk pads with their terminal prolegs and hanging by silk girdles](Battus_polydamas13.jpg)
Shortly before emergence, the patterns of the adult wings and body show through the pupal exoskeleton (Figure 14).
Figure 14. Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]). Preadult with wing and body patterns showing through pupal exoskeleton. Photograph by Donald W. Hall, University of Florida.
![Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]). Preadult with wing and body patterns showing through pupal exoskeleton](Battus_polydamas14.jpg)
Adults usually emerge in the morning and are ready to fly by afternoon (Brown et al. 1980[81], Moss 1919). After emergence, the adult remains near the pupation site while its wings expand and dry (Figure 15). During this time, it also voids the reddish, liquid meconium (waste products from the breakdown of old larval tissues during the pupal stage) from its anus.
Figure 15. Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]). Recently emerged adult drying its wings. Photograph by Donald W. Hall, University of Florida.
![Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]). Recently emerged adult drying its wings](Battus_polydamas15.jpg)
Comstock and Grimshawe (1935) give the following developmental times:
Egg stage: four to six days – average five days at 80 degrees
Larval stage: 19-24 days – average 24 days at 78 degrees and 19 days at 86 degrees. (Early instars molt approximately every four days)
Pupal stage: average 18 days (temperature not specified)
Host Plants (Back to Top)
Larval host plants: Troidine swallowtail larvae feed exclusively on plants belonging to the genus Aristolochia (Aristolochiaceae) (Urzúa & Priestap 1985) often called Dutchman’s pipes or simply pipevines because the flowers of some species are shaped like tobacco pipes (Figure 16).
Figure 16. Unopened flower bud of woolly pipevine, Aristolochia tomentosa Sims, and tobacco pipe graphic showing the similarity of shapes. Photograph of flower bud by Donald W. Hall, University of Florida. Tobacco pipe graphic from PNG Clip Art (graphic modified and pasted into photograph by Donald W. Hall, University of Florida).

Battus polydamas larvae can likely develop on most (if not all) of the Neotropical and temperate Aristolochia species (Lambremont 1954, Masters 1967, Mega et al. 2015, Minno et al. 2005, Scott 1986, Weintraub 1995). However, local populations may differ in how well they develop on a given Aristolochia species (Rios et al. 2016).
Uses of Aristolochia species in traditional medicine: Aristolochia species are also known as birthworts (“wort” is Old English for herbaceous plant) because of their historic use in child birth. The name Aristolochia is from the Greek roots “aristos” (best) and “lochia” (delivery or child birth) (Crosswhite & Crosswhite 1985, Flora of North America (Accessed December 4, 2016)). All members of the family Aristolochiaceae are believed to contain the pharmacologically active aristolochic acids (Chen & Zhu 1987).
Aristolochic acids in herbal medicine products have been implicated as causative agents of renal toxicity and also may be carcinogenic (Heinrich et al. 2009, Schaneberg et al. 2002). An extract of the southwestern pipevine, Aristolochia watsonii Wooton & Standl., was the main ingredient in the snakeroot oil sold by traveling “snakeroot doctors” at medicine shows in the Old West during the 19th century (Crosswhite & Crosswhite 1985).
Although they are now officially banned in many countries, Aristolochia-derived herbal products or parts of the plants themselves are still used in many areas of the world to treat various conditions including snake bite, gastrointestinal problems, respiratory problems, wounds, infectious diseases, and fever (Chen & Zhu 1987, Schaneberg et al. 2002). Aristolochia serpentaria L. (Figure 17), which ranges from central Florida northward in much of the eastern United States, has been used for many medical applications (Heinrich et al. 2009, Moerman 1998).
Figure 17. Virginia snake root (Aristolochia serpentaria L.), a host of the polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]) larva. Photograph by Donald W. Hall, University of Florida.
![Virginia snake root (Aristolochia serpentaria L.), a host of the polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]) larva](Battus_polydamas17.jpg)
The host for the Polydamas swallowtail in the Florida Keys is probably Marsh's Dutchman's pipe, Aristolochia pentandra Jacq. (Comstock & Grimshawe 1935, Minno & Emmel 1993). In New Orleans (von Reisenstein 1863) and Arkansas (Masters 1967), larvae have been recorded from Aristolochia serpentaria.
The native pipevines, woolly pipevine (Aristolochia tomentosa Sims) (Figure 18) and Dutchman’s pipe (Aristolochia macrophylla Lam. – synonyms: Aristolochia durior Hill and Aristolochia sipho L'Heritier, Stirp.) (Figure 19), are sometimes planted in Florida (south of their native ranges) and are used as hosts by Battus polydamas larvae.
Figure 18. Woolly pipevine (Aristolochia tomentosa Sims), a host of the Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]) caterpillar. a) foliage b) unopened flower bud c) opened flower d) longitudinal section through flower showing internal structure. Photographs by Donald W. Hall, University of Florida.
Figure 19. Dutchman’s pipe (Aristolochia macrophylla Lam. [synonyms: Aristolochia durior Hill and Aristolochia sipho L'Heritier, Stirp.]), a host of the Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]) caterpillar. Photographed at Cascade Falls trail, Giles County, Virginia. Photograph by Donald W. Hall, University of Florida.
![Woolly pipevine (Aristolochia tomentosa Sims), a host of the Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]) caterpillar. a) foliage b) unopened flower bud c) opened flower d) longitudinal section through flower showing internal structure](Battus_polydamas18.jpg)
![Dutchman’s pipe (Aristolochia macrophylla Lam. [synonyms: Aristolochia durior Hill and Aristolochia sipho L'Heritier, Stirp.]), a host of the Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]) caterpillar](Battus_polydamas19.jpg)
In addition to native species, various exotic ornamental pipevines (e.g., elegant Dutchman's pipe or calico flower, Aristolochia elegans M.T. Mast [synonym: Aristolochia littoralis Parodi]) (Figure 20) and Aristolochia gigantea Mart. & Zucc. (Figure 21) are also cultured in Florida and are commonly used by Battus polydamas larvae for hosts. Often the larvae become pests by defoliating the vines and eating the flowers. Unfortunately, Aristolochia elegans (and possibly some of the other exotic pipevines) may be a death trap for our other native troidine swallowtail, Battus philenor (L.). Female Battus philenor readily oviposit on Aristolochia elegans, but most of the larvae cannot survive on it (Scott 1986, Tyler 1975). Therefore, planting of Aristolochia elegans is not recommended where Battus philenor occurs (central Florida and northward).
Figure 20. Elegant Dutchman's pipe or calico flower, Aristolochia elegans M.T. Mast [synonym: Aristolochia littoralis Parodi]), a host of the Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]) caterpillar. Photograph by Donald W. Hall, University of Florida; drawing of longitudinal section of flower by Margo Duncan.
Figure 21. Aristolochia gigantea Mart. & Zucc., a host of the Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]) caterpillar. Photograph by Donald W. Hall, University of Florida. (Flowers of the Brazilian form of Aristolochia gigantea have long lobes [Minno et al. 2005])
![Elegant Dutchman's pipe or calico flower, Aristolochia elegans M.T. Mast [synonym: Aristolochia littoralis Parodi]), a host of the Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]) caterpillar](Battus_polydamas20.jpg)
![Aristolochia gigantea Mart. & Zucc., a host of the Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]) caterpillar](Battus_polydamas21.jpg)
Aristolochia pentandra, Aristolochia tomentosa, and Aristolochia macrophylla are vines (Pfeifer 1966) with flattened seeds. The seeds are probably dispersed by wind and possibly also by flowing water and animals. Two Russian Aristolochia vines with similar seeds that grow in similar habitats have been shown to be primarily dispersed by wind and water and to a lesser extent by birds (Nechaev & Nakonechnaya 2009). Aristolochia serpentaria is a small herb. Its seeds have an attached oil body (elaiosome) and are dispersed by ants which collect the seeds for the edible elaiosomes (D.W. Hall, personal observation).
Aristolochia species have fascinating pollination biologies (Proctor et al. 1996). When the flowers first open, the stigmas are receptive, but the anthers are not releasing pollen – a phenomenon known as protogyny (from the Greek roots proto [first] and gyne [female]). During the first day of bloom, the flowers are attractive usually to small flies (often but not always scuttle flies in the family Phoridae [Hall & Brown 1993]) which enter the flowers carrying pollen from another flower. The flies are then prevented from leaving by the presence of downward projecting guard hairs or tiny spines and slippery surfaces in the tube of the flower (Proctor et al. 1996, Zomlefer 1994) (Figures 18 & 20).
On the second day (after pollination), the stigmas become non-receptive, and the anthers dehisce (open) releasing pollen which becomes attached to the flies. The guard hairs in the flower tubes wither (and in some species [e.g., Aristolochia grandiflora] the flowers also droop into a more horizontal position [Burgess et al. 2004]) releasing the flies that are now carrying the new pollen.
Nectar host plants: Battus polydamas adults feed on nectar from a variety of flowers and are reported to be particularly fond of the flowers of the common exotic weed Lantana camara L. (Opler & Krizek 1984) (Figure 22).
The new flowers of Lantana camara are in the center of the flower cluster. They are lighter in color (shades of yellow) and produce nectar. After pollination, they turn a darker color (pink or red), stop producing nectar and flower volatiles, and the stigmas become unreceptive (Raguso & Weiss 2015, Ram & Mathur 1984, Weiss 1991, Weiss 1997). Butterflies learn to visit only the lighter colored, nectar-producing flowers. However, the old flowers (now on the outer rings of the cluster) remain on the plant – possibly to make the larger flower cluster more attractive to new pollinators and to provide a landing platform for larger pollinators (e.g., butterflies).
Figure 22. Orange (top) and pink (bottom) varieties of the invasive exotic Lantana camara L., a favored nectar host plant for the Polydamas swallowtail, Battus polydamas lucayus (Rothschild & Jordan). Note new, light-colored flowers on interior of clusters. Photographs by Donald W. Hall, University of Florida.

Lantana camara is listed as a Category 1 invasive on the Florida Exotic Pest Plant Council’s 2015 List of Invasive Plant Species (Accessed 23 March 2020). Therefore, this species should not be planted.
Flower Gardens. Minno and Minno (1999) have extensive lists of both native and exotic nectar plants for butterflies. When possible, native plants should be planted as nectar sources rather than exotics that have the potential to be invasive.
Most states have native plant societies that are valuable sources of information on native plants and many also hold native plant sales.
For Florida and the Deep South, the Florida Wildflowers Growers Cooperative (http://www.floridawildflowers.com/) (Accessed 23 March 2020) is an excellent source of information and also has wildflower seeds for purchase.
To maximize butterfly populations in yards and gardens, both caterpillar hosts and nectar plants for adults should be planted. The choice of which one(s) to plant is dependent on locality.
Medicinal Uses (Back to Top)
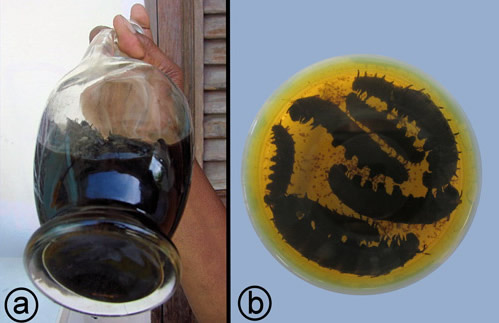
Larvae of the Aristolochia swallowtails sequester aristolochic acids and secondary metabolites from their host plants (Priestap et al. 2012). An alcoholic extract (usually in rum) of Battus polydamas larvae (that have fed on Aristolochia trilobata L.) has been used as a medicinal product known as Chiniy-trèf on the Caribbean island of Martinique (Cachet et al. 2016, Nossin & Weniger 2008) (Figure 23) but its sale is now prohibited.
Figure 23. Chiniy-trèf, an alcoholic extract of Battus polydamas larvae. a) batch preparation. Photograph by Emmanuel Nossin, Hôpital du Lamentin, Service Pharmacie, Martinique. b) sample, showing larvae. Photograph by Cécile Bottai, Faculty of Pharmacy, Paris Descartes University, Paris, France.
Defenses (Back to Top)
Eggs: Young (1973), in a discussion of another Aristolochia swallowtail (Parides arcas mylotes Bates), stated that it is unknown whether the secretion on the outside of the eggs may be defensive against ants and other potential predators. The fact that the brightly colored secretion (Figure 24) makes the eggs conspicuous suggests a possible aposematic (warning) function.
Figure 24. Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]). Eggs showing detail of brightly colored secretion. Photograph by Donald W. Hall, University of Florida.
![Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]). Eggs showing detail of brightly colored secretion](Battus_polydamas24.jpg)
Larvae, pupae and adults: Larvae of the Aristolochia swallowtails sequester aristolochic acids from their host plants, and concentrations in their tissues are probably due to the relative concentrations in the host plants (Urzúa & Priestap 1985). Also, the aristolochic acids (and possibly other Aristolochia phytochemicals [Morais et al. 2013]) render larvae, pupae, and adults unpalatable to many natural enemies. This chemical defense is enhanced in adults by their bright aposematic (warning) coloration. Pupae resemble green or brown leaves and likely gain more protection from their cryptic coloration than from chemical defenses.
In laboratory studies, Battus polydamas larvae were repellent to domestic chickens (Gallus gallus domesticus [L.]) and the carpenter ant Camponotus crassus Mayr but not to the assassin bug Montina confusa Stål (Morais et al. 2013). In a field study, adult mortality of Battus polydamas was low - a result attributed to low predation rates by birds (Young 1972).
Adults of two other Aristolochia swallowtails, Battus philenor and Parides neophilus (Geyer), have been shown to be chemically defended against caged birds (Brower 1958, Brower & Brower 1962, Brower & Brower 1964).
All swallowtail larvae have eversible organs (osmeteria) on the anterior margin of the prothorax. The osmeterium of the Polydamas swallowtail is bright orange (Figure 25). The bright coloration of the osmeterium may startle potential predators. The osmeterium was ineffective against the attack of an ovipositing ichneumonid wasp, Areoscelis rufa Brullé (Tyler et al. 1994, Plate 6).
Figure 25. Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]). Larva with osmeterium extruded. Photograph by Donald W. Hall, University of Florida.

The osmeterial secretions of all other swallowtails that have been examined (species belonging to the tribes Graphiini and Papilionini of the subfamily Papilioninae and to the subfamily Baroniinae) are composed of isobutyric and 2-methylbutyric acids (Eisner et al. 1971). However, Battus polydamas (Papilioninae: Troiidini) secretes the two sesquiterpenes selin-11-en-4α-ol and β-selinene (Eisner et al. 1971) in addition to aristolochic acids (Priestap et al. 2012).
Prepupae retain the ability to evert the osmeteria right up to the time of pupation (Figure 26).
Figure 26. Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]). Prepupa with osmeterium extruded (Note silk girdle [arrow] suspending prepupa). Photograph by Donald W. Hall, University of Florida.
![Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]). Prepupa with osmeterium extruded (Note silk girdle [arrow] suspending prepupa)](Battus_polydamas26.jpg)
The reddish-orange bands on the abdomen of adults (Figure 27) contain glands that secrete an acrid odor when the butterflies are pinched (Tyler 1975, p.155).
Figure 27. Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]) showing reddish-orange bands on abdomen. Photograph by Donald W. Hall, University of Florida.
![Polydamas swallowtail (Battus polydamas lucayus [Rothschild & Jordan]) showing red spots on abdomen.](Battus_polydamas27.jpg)
Acknowledgements (Back to Top)
The author would like to acknowledge Marc Minno and Howard Frank for reviewing this article and offering helpful suggestions and Emmanuel Nossin and Cécile Bottai for permission to use their photographs of Chiniy-trèf preparations.
Selected References (Back to Top)
- Barbier R, Chauvin G. 1976. L’enveloppe vitelline chez les Lépidoptères: transformation lors de l’activation de l’ovocyte et liaison avec la cuticule sérosale. Bulletin de la Sociètè Zoologique de France. 101: 1084-1085.
- Brown KS, Damman AJ, Feeny P. 1980(81). Troidine swallowtails (Lepidoptera: Papilionidae) in southeastern Brazil: natural history and foodplant relationships. Journal of Research on the Lepidoptera 19: 199-226.
- Brower JV. 1958. Experimental studies of mimicry in some North American butterflies. 2. Battus philenor and Papilio troilus, P. polyxenes and P. glaucus. Evolution 12: 123-136.
- Brower LP, Brower JVZ. 1962. The relative abundance of model and mimic butterflies in natural populations of the Battus philenor mimicry complex. Ecology 43: 154-158.
- Brower LP, Brower JVZ. 1964. Birds, butterflies, and plant poisons: a study in ecological chemistry. Zoologica (New York) 49: 137-159.
- Burgess KS, Singfield J, Melendez V, Kevan PG. 2004. Pollination biology of Aristolochia grandiflora (Aristolochiaceae) in Veracruz, Mexico. Annals of the Missouri Botanical Garden 91: 346-356.
- Cachet X, Langrand J, Bottai C, Dufat H, Locatelli-Jouians C, Nossin E, Boucaud-Maitre D. 2016. Detection of aristolochic acids I and II in “Chiniy-trèf”, a traditional medicinal preparation containing caterpillars feeding on Aristolochia trilobata L. in Martinique, French West Indies. Toxicon 114: 28-30.
- Chen Z-L, Zhu D-Y. 1987. Aristolochia Alkaloids. Chap. 2. In: Brossi A, ed. The Alkaloids: Chemistry and Pharmacology. Academic Press. New York. pp. 29-65.
- Comstock JA, Grimshawe FM. 1935. Early stages of Papilio polydamas lucayus R. & J. Bulletin of the Southern California Academy of Sciences 34: 76-80
- Crosswhite FS, Crosswhite CD. 1985. The southwestern pipevine (Aristolochia watsonii) in relation to snakeroot oil, swallowtail butterflies, and ceratopogonid flies. Desert Plants 6: 203-207.
- dos Passos CF. 1940. On the occurrence of Papilio polydamas Linnaeus within the United States. The Canadian Entomologist 72: 188.
- Eisner T, Kluge AF, Ikeda MI, Meinwald YC, Meinwald J. 1971. Sesquiterpenes in the osmeterial secretion of a papilionid butterfly, Battus polydamas. Journal of Insect Physiology 17: 245-250.
- Field WD. 1938. A manual of the butterflies and skippers of Kansas (Lepidoptera: Rhopalocera). Bulletin of the University of Kansas: Biological Series.39: p.199.
- Hall DW, Brown BV. 1993. Pollination of Aristolochia littoralis (Aristolochiales: Aristolochiaceae) by males of Megaselia spp. (Diptera: Phoridae). Annals of the Entomological Society of America 86: 609-613.
- Heinrich M, Chan J, Wanke S, Neinhuis C, Simmonds MSJ. 2009. Local uses of Aristolochia species and content of nephrotoxic aristolochic acid 1 and 2 - a global assessment based on bibliographic sources. Journal of Ethnopharmacology 125: 108-144.
- Lambremont EN. 1954. The butterflies and skippers of Louisiana. Tulane Studies in Zoology 1: 125-164.
- Lindsey AW. 1925. The types of hesperioid genera. Annals of the Entomological Society of America 18: 75-106.
- Masters JH. 1967. Observations on Arkansas Rhopalocera and a list of species occurring in northeastern Arkansas. Journal of the Lepidopterists’ Society 21: 206-209.
- Mega NO, Scalco VW, Atencio GWG, Morais ABB, Romanowski HP. 2015. Battus polydamas (Lepidoptera: Papilionidae) uses the open-field Aristolochia sessilifolia (Piperales: Aristolochiaceae) as its host plant in Uruguayan savanna areas. Florida Entomologist 98: 762-769.
- Miller JS. 1987. Phylogenetic studies in the papilioninae (Lepidoptera: Papilionidae). Bulletin of the American Museum of Natural History 186: 365-512.
- Minno MC, Butler JF, Hall DW. 2005. Florida Butterfly Caterpillars and their Host Plants. University Press of Florida. Gainesville, Florida. 341 pp.
- Minno MC, Emmel, TC. 1993. Butterflies of the Florida Keys. Scientific Publishers. Gainesville, Florida. 168 pp.
- Minno MC, Minno M. 1999. Florida Butterfly Gardening. University Press of Florida. Gainesville, Florida. 210 pp.
- Moerman. DE. 2009. Native American Ethnobotany. Timber Press. Portland, Oregon. 927 pp.
- Morais ABB, Brown KS Jr, Stanton MA, Massuda KF, Trigo JR. 2013. Are aristolochic acids responsible for the chemical defense of aposematic larvae of Battus polydamas (L.) (Lepidoptera: Papilionidae)? Neotropical Entomology 42: 558-564.
- Moss AM. 1919. The papilios of Para. Novitates Zoologicae 26: 295-319.
Nechaev VA, Nakonechnaya OV. 2009. Structure of fruits and seeds and ways of dissemination of two species of the genus Aristolochia L. in Primorsky Krai. Biology Bulletin 36: 393-396.
- Nossin E, Weniger B. 2008. Sur Aristolochia trilobata et ses usages traditionnels en Martinique et dans la Caraibe. Ethnopharmacologia No. 42. (December). pp. 37-43.
- Opler PA, Krizek GO. 1984. Butterflies East of the Great Plains. The Johns Hopkins University Press. Baltimore, MD. 294 pp.
- Opler PA, Malikul V. 1998. Eastern Butterflies. Peterson Field Guide Series. Houghton Mifflin. New York. 486 pp.
- Pfeifer HW. 1966. Revision of the North and Central American hexandrous species of Aristolochia (Aristolochiaceae). Annals of the Missouri Botanical Garden 53: 115-196.
- Pinto CF, Troncoso AJ, Urzúa A, Niemeyer M. 2009a. Aristolochic acids affect the feeding behavior and development of Battus polydamas archidamas larvae (Lepidoptera: Papilionidae: Troidini). European Journal of Entomology 106: 357-361.
- Pinto CF, Troncoso AJ, Urzúa A, Niemeyer M. 2009b. Use of volatiles of Aristolochia chilensis (Aristolochiaceae) in host searching by fourth-instar larvae and adults of Battus polydamas archidamas (Lepidoptera: Papilionidae: Troidini). European Journal of Entomology 106: 63-68.
- Priestap HA, Velandia AE, Johnson JV, Barbieri MA. 2012. Secondary metabolite uptake by the Aristolochia-feeding papilionoid butterfly Battus polydamas. Biochemical Systematics and Ecology 40: 126-137.
- Proctor M, Yeo P, Lack A. 1996. The Natural History of Pollination. Timber Press. Portland, Oregon. 479 pp.
Racheli T, Oliverio M. 1993. Biogeographical patterns of the neotropical genus Battus Scopoli 1777 (Lepidoptera: Papilionidae). Tropical Zoology 6: 55-65.
- Raguso RA, Weiss MR. 2015. Concerted changes in floral colour and scent and the importance of spatio-temporal variation in floral volatiles. Journal of the Indian Institute of Science 95: 69-92.
- Ram H, Mathur G. 1984. Flower color changes in Lantana camara. Journal of Experimental Botany 35: 1656-1662.
- Rausher MD. 1979. Larval habitat suitability and oviposition preference in three related butterflies. Ecology 60: 503-511.
- Richards OW, Davies RG. 1977. Chap. 18. Embryology. In: Vol. 1. Imms’ Textbook of Entomology. 10th edition. John Wiley & Sons. New York. 418 pp.
- Rios RS, Salgado-Luarte C, Stotz GC, Gianoli E. 2016. Co-occurrence of host plants associated with plant quality determines performance patterns of the specialist butterfly, Battus polydamas archidamas (Lepidoptera: Papilionidae: Troidini). European Journal of Entomology 113: 150-157.
- Rothschild W, Jordan K. 1906. A revision of the American Papilios. Novitates Zoologicae 13(3): 411-752. http://www.biodiversitylibrary.org/item/21926#page/425/mode/1up (4 December 2016)
- Schaneberg BT, Applequist WL, Khan IA. 2002. Determination of aristolochic acid I and II in North American species of Asarum and Aristolochia. Pharmazie 57: 686-689.
- Scopoli, G.A. 1777. Introductio ad historiam naturalem sistens genera lapidum, plantarum, et animalium :hactenus detecta, caracteribus essentialibus donata, in tribus divisa, subinde ad leges naturae. Apud Wolfgangum Gerle. Prague. p. 433. http://www.biodiversitylibrary.org/item/42076#page/1/mode/1up (23 March 2020)
- Scott JA. 1986. The Butterflies of North America. Stanford University Press. Stanford, California. 583 pp.
Spade P, Tyler H, Brown JW. 1988. The biology of seven troidine swallowtail butterflies (Papilionidae) in Colima Mexico. Journal of Research on the Lepidoptera 26: 13-26.
- Tyler HA. 1975. The Swallowtail Butterflies of North America. Naturegraph Publishers. Healdsburg, California. 192 pp.
- Tyler H, Brown KS Jr., Wilson K. 1994. Swallowtail Butterflies of the Americas: A Study in Biological Dynamics, Ecological Diversity, Biosystematics, and Conservation. Scientific Publishers. Gainesville, Florida. 376 pp.
- Urzúa A, Priestap H. 1985. Aristolochic acids from Battus polydamas. Biochemical Systematics and Ecology 13: 169-170.
- von Reizenstein L. 1863. Catalogue of the Lepidoptera of New Orleans and its vicinity. Isaac T. Hinton. New Orleans. pp. 1-8. (Original not seen - Cited by Lambremont EN. 1954)
- Weintraub JD. 1995. Host plant association patterns and phylogeny in the tribe Troidini (Lepidoptera: Papilionidae). Chap. 27. In: Scriber JM, Tsubaki Y, Lederhouse RC. Swallowtail Butterflies: Their Ecology & Evolutionary Biology. Scientific Publishers. Gainesville, Florida. 459 pp.
- Weiss MR. 1991. Floral colour changes as cues for pollinators. Nature 354: 227-229.
- Weiss MR. 1997. Innate colour preferences and flexible colour learning in the pipevine swallowtail. Animal Behaviour 53: 1043-1052.
- Young AM. 1972. Mimetic associations in populations of tropical butterflies. II. Mimetic interactions of Battus polydamus and Baltus bellus.Biotropica 4: 17-27.
- Young AM. 1973. Notes on the life cycle and natural history of Parides arcas mylotes (Papilionidae) in Costa Rican premontane wet forest. Psyche 80: 1-21.
- Zomlefer WB. 1994. Plant Families. The University of North Carolina Press. Chapel Hill, North Carolina. 430 pp.